Structural insight into M-band assembly and mechanics from the titinobscurin- like-1 complex
Nov 5, 2010
Nov 5, 2010
Dr Roberto Steiner, King’s College London
In muscle, assembly of ordered striated myofibrils requires the integration of hundreds of protein subunits in a sequential, regulated way by specific protein-protein interactions. The contractile filaments, composed of actin and myosin subunits, are crosslinked at the Z-disk and M-band, respectively, to give rise to a lateral assembly of molecular machines with nanometre precision. In the sarcomeric M-band, the giant ruler proteins titin and obscurin, its small homologue obscurin-like-1 (Obsl1), and the myosin cross-linking protein myomesin form a ternary complex that is crucial for the function of the M-band as a mechanical link. Mutations in the last titin immunoglobulin (Ig) domain M10, which interacts with the N-terminal Ig-domains of obscurin and Obsl1, lead to hereditary muscle diseases. The M10 domain is unusual not only in that it is a frequent target of disease-linked mutations, but also in that it is the only currently known muscle Ig-domain that interacts with two ligands—obscurin and Obsl1—in different sarcomeric sub-regions. Using X-ray crystallography complemented by pull-down assays, isothermal titration calorimetry, single-molecule force-microscopy and cellular competition assays in cardiomyocytes, we show the structural basis for titin M10 interaction with Obsl1 in a novel antiparallel Ig-Ig architecture and unravel the molecular basis of titin-M10 linked myopathies. The severity of these pathologies correlates with the disruption of the titin-obsl1/obscurin complex. Conserved signature residues at the interface account for differences in affinity that direct the cellular sorting in cardiomyocytes. By engineering the interface signature residues of obsl1 to obscurin, and vice versa, their affinity for titin can be modulated similar to the native proteins. In single-molecule force-spectroscopy experiments, both complexes yield at forces of around 30 pN, much lower than those observed for the mechanically stable Z-disk complex of titin and telethonin, suggesting why even moderate weakening of the Obsl1/obscurintitin links has severe consequences for normal muscle functions.
Sarcomeres are the smallest contractile units of striated muscles. They are highly ordered assemblies of precisely tailored actin and myosin filaments that are crosslinked at Z-disks and M-bands, respectively. The assembly of hundreds of protein subunits into ordered sarcomeres forms the structural basis for striated muscle contraction. The global layout of sarcomere assembly requires a giant ruler protein, titin, a 3000 kDa modular protein with a length of over 1.2 µm, displaying binding sites for proteins along the entire distance from Z-disk to M-band [1]. Titin is composed of hundreds of immunoglobulin- and fibronectin-3- like (Ig and Fn3) domains that are arranged in specific patterns in the sub-compartments of the sarcomere. The Ig-domains in sarcomeric proteins like titin, myomesin or obscurin present a functionally versatile surface around a highly stable structural scaffold [2]. At the M-band, the centers of myosin filaments are crosslinked into a mechanically stable network that involves titin, the cytoskeletal protein myomesin, and the giant obscurin, the latter acting as a linker to the sarcoplasmic reticulum (SR). Obscurin, with a mass of around 800 kDa, was initially discovered as a ligand of Z-disk titin [1]. Like titin, it is a modular protein composed of Ig and Fn3 domains. Intriguingly, obscurin localises predominantly to the M-band in mature myofibrils, a behaviour unexpected for a giant sarcomeric protein. This multi-compartment localization is explained by the presence of a further, M-band-specific binding site for obscurin at the most C-terminal titin domain M10. This domain interacts with the N-terminal domain of both obscurin and its small homologue, obscurin-like-1 (Obsl1), which lacks the signalling domains and ankyrin-binding sites that link the obscurin C-terminus to the SR. In agreement with the SR interaction, obscurin is localised to the myofibril periphery, whereas Obsl1 is an integral component of the myofibril [3]. Titin’s C-terminus plays a critical role in muscle function, as multiple mutations at this position cause hereditary myopathies. Recently, mutations in Obsl1 were identified as the cause of the primordial growth disorder 3-M syndrome.
To understand at the atomic level the sarcomeric M-band interaction between titin and Obsl1, we reconstituted the minimal interacting complex in vitro and solved its X-ray crystal structure by the molecular replacement technique in two different crystal forms (Form I, trigonal and Form II, triclinic – PDB codes 2wp3 and 2wwm, respectively). Data were collected at Diamond Light Source beam lines I04 and I03. We complemented our crystallographic analysis by pull-down assays, isothermal titration calorimetry (ITC), single-molecule force-microscopy and cellular competition assays in cardiomyocytes. Titin M10 (hereafter M10) and the N-terminal Ig module of human Obsl1 (hereafter OL1) form a heterodimer in solution and in the crystal. No major structural differences are seen in the two different crystal forms, suggesting that the complex observed crystallographically faithfully recapitulates that in solution. After restrained refinement, the best M10-OL1 model solved at the 1.48 Å resolution is characterised by an Rfree value of 21.7% and excellent quality indicators. The M10-OL1 heterodimer is shown in Figure 1AB. Both M10 and OL1 protomers belong to the intermediate set (I-set) of the Ig family and share the typical barrel-like architecture in which a total of nine strands are arranged into two distinct ß-sheets folded into a ß-sandwich. A mixture of hydrogen bonds, salt bridges and hydrophobic interactions holds the complex together (Figure 1C). A search in the SCOPPI [4] database of protein-protein interfaces reveals that the M10-OL1 complex represents a mode of interaction previously unobserved for Ig domains. The structure of the M10-OL1 complex reveals that disease-linked residues are not directly involved in the proteinprotein interface. Rather, the predicted outcome of the amino acid variations induced by the Finnish and French mutations is a severe disruption of the M10 fold.
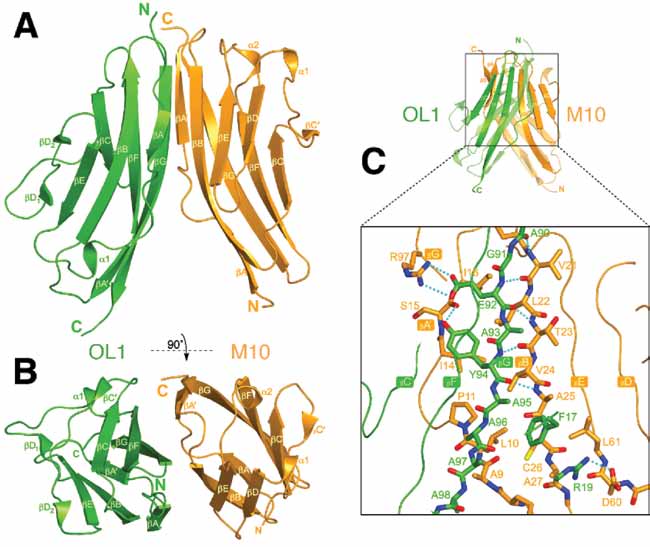 |
| Figure 1. (A and B) Cartoon representation of the M10-OL1 heterodimer. As indicated, the view in (B) is rotated by 90 degrees around the x axis compared to the view in (A). M10 and OL1 are shown in orange and green, respectively. (C) Enlarged view of the boxed overall representation shown in the top panel high-lighting important M10-OL1 interactions. Some structural elements in the overall representation are shown partly transparent to highlight the mixed inter-molecular β-sheet. Colour coding for carbon atoms is orange and green for M10 and OL1, respectively; nitrogen, blue; oxygen, red; sulfur, yellow. Hydrogen bonds are dotted cyan lines. |
ITC measurements indicate that the formation of both M10-OL1 and M10-O1 complexes is driven by entropic gain rather than by enthalpic factors. The M10-OL1 complex is more stable than the M10-O1 complex, with a 3.4-fold higher affinity than the latter. The OL1 and O1 domains are 38.5% identical and display a strong conservation of the residues involved in forming the M10-OL1 interface suggesting that O1 binds to M10 similarly to OL1. A striking difference, however, is the replacement of OL1 F17 with an arginine residue (R15) in O1. OL1 F17 interacts with a hydrophobic pocket on M10 using its aromatic side-chain. F17 and R15 are totally conserved within the vertebrate Obls1 and obscurin families, respectively, effectively representing a signature of their Ig1 domains. To understand in more detail the effect of an arginine residue at this position, we also solved the crystal structure of the M10-OL1 F17R complex (PDB code 2wwk). The structure reveals that “obscurin-ization” of OL1 only affects the complex locally, resulting in the positively charged arginine side-chain pointing towards the solvent. A cluster of three ordered water molecules fills the volume occupied by the aromatic side-chain of F17 in the M10-OL1 complex. In agreement with the thermodynamic data, the decrease in stability due to the OL1 F17R replacement can be linked to a loss of entropic gain. The highly conserved, discriminating surfaces in M10-OL1 and M10-O1 account for specific differences in affinity that are reflected in the cellular specificity of their sarcomeric interactions. That the key residue differences are biologically significant is underscored by their remarkable cross-species conservation. High affinity interaction of titin with Obsl1 is involved in formation of core myofibrils, whereas lower-affinity interaction directs the peripheral integration of obscurin albeit our assays in cardiomyocytes suggests that additional elements of specificity might be involved. The antiparallel orientation of titin and obsl1/obscurin will need to be considered in future revised M-band models.
References
[1] Lange S, Ehler E, Gautel M, Trends Cell. Biol., 16, 11–18 (2006).
[2] Otey CA, Dixon R, Stack C, Goicoechea SM, Cell Motil. Cytoskeleton, 66, 618–634 (2009).
[3] Fukuzawa A, Lange S, Holt M, Vihola A, Carmignac V, Ferreiro A, Udd B, Gautel M, J. Cell Sci., 121, 1841–1851 (2008).
[4] Winter C, Henschel A, Kim WK, Schroeder M, Nucleic Acids Res., 34, D310–314 (2006).
Principal Publications and Authors
Pernigo S, Fukuzawa A, Bertz M, Holt M, Rief M, Steiner RA, Gautel M. Structural insight into M-band assembly and mechanics from the titin-obscurin-like 1 complex. Proc. Natl. Acad. Sci. USA, 107, 2908-2913 (2010).
Funding Acknowledgement
King’s College London, British Heart Foundation (BHF) Centre of Research Excellence and the Medical Research Council, UK.

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Copyright © Diamond Light Source. Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.