Gram-negative bacteria, mitochondria and chloroplasts all have outer membranes, which contain a variety of proteins. As well as helping to pull together the membrane components, outer membrane proteins have an important role in importing nutrients, exporting waste products and transporting proteins. In Gram-negative bacteria, outer membrane proteins are inserted and folded into the outer membrane by the β-barrel assembly machinery (BAM), but the mechanism of this process is poorly understood. BAM itself is a complex that comprises five subunits: one outer membrane protein called BamA and four lipoproteins (fat-loving proteins that are fixed to the membrane) known as BamB, Bam C, BamD and BamE.
To shed some light on how BAM works, two crystal structures were solved using the Macromolecular X-ray Crystallography beamlines (I02, I03 and I24), at Diamond Light Source. The first structure was BAM lacking BamB (BamACDE) and the second structure was Bam in its entirety (BamABCDE).
The crystal structures showed that the BAM subunits assembled into a unique shape resembling a top hat, which was embedded within the outer membrane. Comparing the structures revealed that BAM occupied two distinct conformational states, which were snapshots of the machinery in action. Furthermore, in conjunction with functional studies, it was determined that the BAM subunits rotated to encourage the insertion of proteins into the outer membrane.
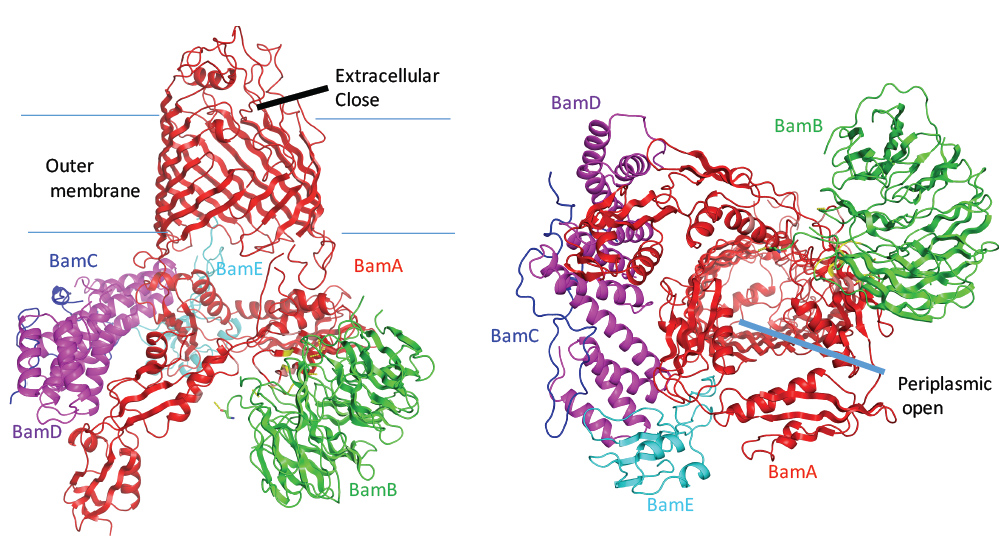
Figure 1: The entire BAM machinery (BamABCDE) structure; (a) The side view of the BAM machinery; (b) The periplasm view of the BAM machinery in the inward-open conformation .
Bacterial drug resistance is becoming a serious global health threat, and millions of deaths are caused by drug-resistant bacterial infections in the world every year1. In the EU, drug-resistant bacteria kill 25,000 people each year, of which two-thirds are caused by drug-resistant Gram-negative bacteria. No novel drug families have been introduced into the market in the past thirty years which are effective against Gram-negative bacteria, whilst the utility of existing drugs is eroded by the increase of resistant strains. Many antibiotics can kill Gram-positive bacteria efficiently but are useless against Gramnegative bacteria as these antibiotics cannot bypass the outer membrane to enter into the Gram-negative bacteria.
The outer membrane (OM) is essential for all Gram-negative bacteria, and plays a critical role in bacterial survival in different environments2. The OM is composed of lipopolysaccharide in the outer leaflet and phospholipids in the inner leaflets, which form a barrier that prevents molecular exchanges between the bacteria and the environments. OM proteins (OMPs) are responsible for Gram-negative bacterial nutrient import, waste export, protein translocation, bacterial adhesion, invasion and immune evasion, which are critical for pathogenesis and drug resistance. In E.coli, the OMPs are synthesized in the cytoplasm, and translocated across the inner membrane (IM) into the periplasm by SecYEG, where chaperon proteins, such as SurA and Skp, escort the unfolded OMPs across the periplasm to the outer membrane barrel assembly machinery (BAM). The BAM machinery contains one OMP BamA and four lipoproteins, BamB, BamC, BamD and BamE, in which BamA and BamD are essential in E.coli, and the BAM machinery is reported as a novel drug target. However, the mechanism of how the BAM machinery inserts and assembly outer membrane proteins is unknown3. Additionally, OMPs are also critical in outer membrane biogenesis in mitochondria and chloroplasts, where OMP85 family proteins are responsible for the outer membrane protein insertion4. The OMP85 proteins are predicted to form 16 b-stranded barrels as BamA. Therefore, the structural study of the BAM machinery is certainly important for understanding the outer membrane biogenesis of mitochondria and chloroplasts.
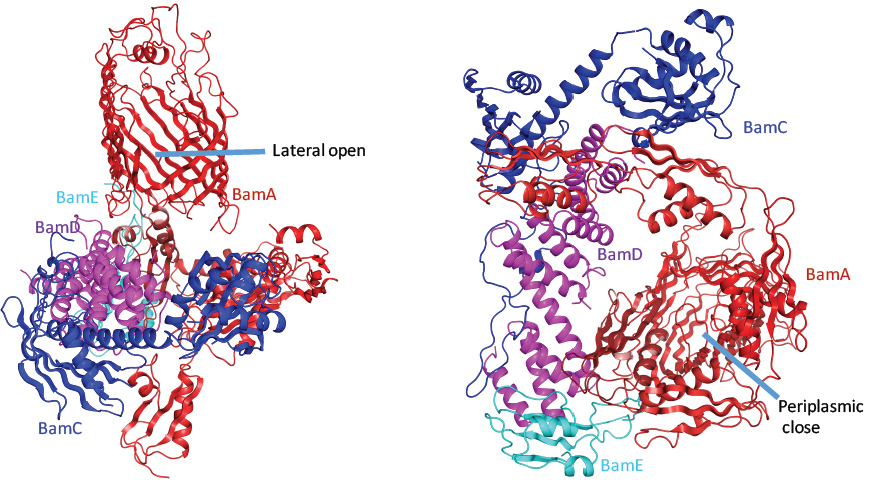
Figure 2: The four-protein BamACDE structure; (a) The side view of the BamACDE complex; (b) The periplasm view of the BamACDE structure in the lateral-open conformation.
BamA is an outer membrane protein, consisting of a carboxyl-terminal 16 b-stranded membrane barrel and five amino-terminal polypeptide transport associated (POTRA) domains. The BamABCDE complex was crystallised and the structure was determined. BamA POTRA domains and lipoprotein BamD form a periplasmic ring, where BamA POTRA domain 5 interacts with the C-terminal domain of BamD, and BamD’ N-terminal domain interacts with POTRA domain 1 and 2. BamB binds between POTRA domain 2 and 3, and BamC binds BamD and POTRA1 and 2. In addition, BamE is located between BamA and BamD, and interacts with BamC. In the BamABCDE complex, the barrel of the BamA is closed at the extracellular side and is widely opened to the periplasmic side (Fig. 1). Functional assays showed that the residues involved in the POTRA domain 5 and the BamD interactions (BamA residues Arg366, Glu373 and BamD residue Arg197) are critical for the cell growth.
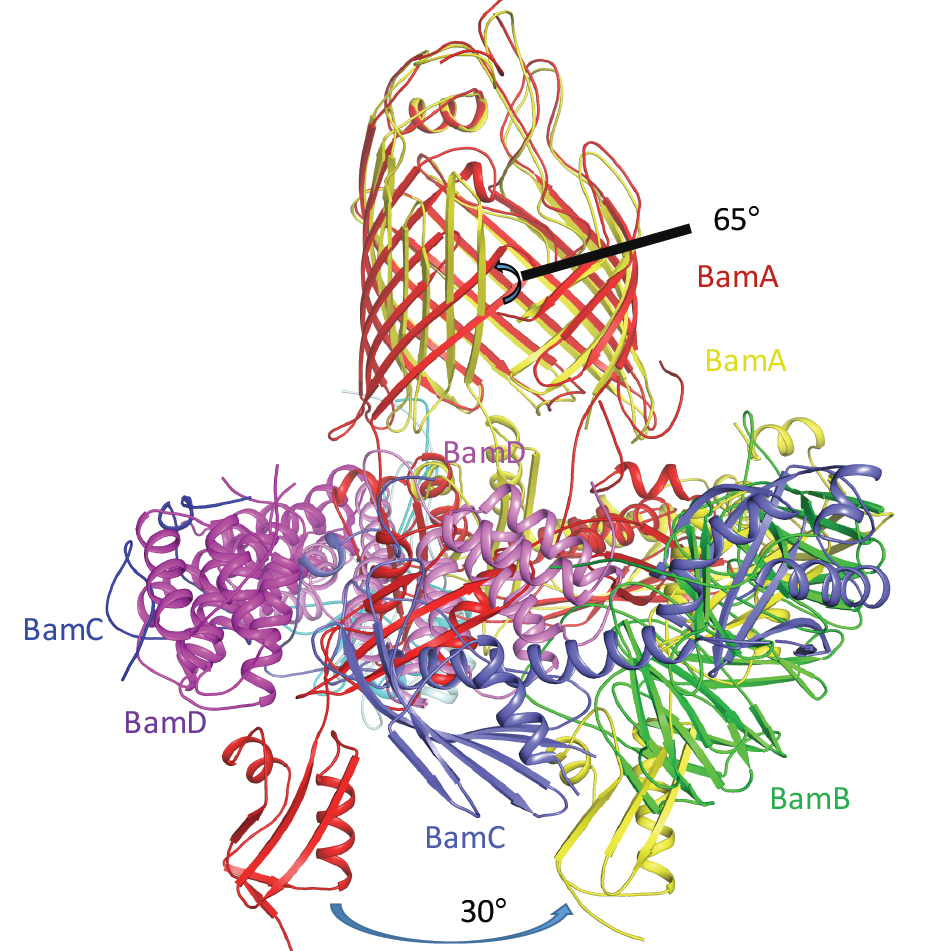
Figure 3: The conformational changes between the two BAM complexes. The two complexes are superimposed at the barrel sections.
BamACDE complex was also crystallised and the structure was determined, which showed that the structure of BamACDE is of a totally different conformation than that of BamABCDE. The extracellular side of BamA barrel is opened, as well as the lateral gate, while the periplasmic side is closed, where the BamA is at a lateral opening conformation (Fig. 2). To confirm whether the two conformations exist in the bacteria, two double cysteine mutants E435C/S665C and E435C/S658C were generated to lock the BAM machinery in the inward-open conformation, and a double cysteine mutant Gly393Cys/ Gly584Cys to lock the BAM machinery in the lateral-open conformation. The functional assays showed that all three mutants are lethal, but could be rescued by 2 mM reducing reagent Tris(2-carboxyethyl)phosphine hydrochloride that break the disulfide bonds, indicating that the two conformations exist in bacteria.
Superimposing the two structures, the C-terminal barrel b1-b6 have significant conformational changes, where the strands rotate around 65° at their central points and shift about 15 Å. At the same time the periplasmic ring rotates about 30° (Fig. 3)
The data suggest that the unfolded outer membrane proteins would interact with the periplasmic ring and enter into the periplasmic ring, triggering the periplasmic ring rotation and promote the BamA barrel strands b1-b6 movement, which will insert the outer membrane proteins into the outer membrane.
References:
- Brown, E. D. et al. Antibacterial drug discovery in the resistance era. Nature. 529(7586), 336-343, doi:10.1038/nature17042 (2016).
- Raetz, C. R. et al. Lipopolysaccharide endotoxins. Annu Rev Biochem. 71, 635-700, doi:10.1146/annurev.biochem.71.110601.135414 (2002).
- Noinaj, N. et al. The β-barrel membrane protein insertase machinery from Gram-negative bacteria. Curr. Opin. Struct. Biol. 31, 35-42, doi.org/10.1016/j.sbi.2015.02.012 (2015).
- Walther, D. M. et al. Biogenesis of β-barrel membrane proteins in bacteria and eukaryotes: evolutionary conservation and divergence. Cell. Mol. Life Sci. 66, 2789–2804, doi:10.1007/s00018-009-0029-z (2009).
Funding acknowledgement:
This work was supported by Wellcome Trust (WT106121MA) to C.J. D.
Corresponding author:
Professor Changjiang Dong, Norwich Medical School, University of East Anglia, [email protected]
Related publication:
Gu Y, Li H, Dong H, Zeng Y, Zhang Z, Paterson NG, Stansfeld PJ, Wang Z, Zhang Y, Wang W, Dong C. Structural basis of outer membrane protein insertion by the BAM complex. Nature 531(7592), 64-69, doi:10.1038/nature17199 (2016).
Publication keywords:
Gram-negative bacteria; β-barrel assembly machinery (BAM); Membrane proteins; X-ray crystallography

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Copyright © Diamond Light Source. Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.