A step towards earlier detection of multiple sclerosis
Aug 9, 2019
Aug 9, 2019
Related publications: Shaharabani R., Ram-On M., Talmon Y. & Beck R. Pathological transitions in myelin membranes driven by environmental and multiple sclerosis conditions. Proc. Natl. Acad. Sci. 115, 11156–11161 (2018). DOI: 10.1073/pnas.1804275115; Shaharabani R., Ram-On M., Avinery R., Aharoni R., Arnon R., Talmon Y. & Beck R. Structural Transition in Myelin Membrane as Initiator of Multiple Sclerosis. J. Am. Chem. Soc. 138, 12159–12165 (2016). DOI: 10.1021/jacs.6b04826
Publication keywords: SAXS; Phase transition; Myelin; Multiple sclerosis; Lamellar phase; Inverted hexagonal phase; Membrane; Lipids
Using Diamond Light Source’s Small Angle Scattering & Diffraction beamline (I22), I911-SAXS at MAX IV Laboratory, SWING at SOLEIL synchrotron, and P12 at EMBL BioSAXS, a team of researchers from Tel Aviv University and the Technion-Israel Institute of Technology mapped, for the first time, the delicate and complicated force balance between the myelin sheath constituents and their effect on the myelin structure.
It is well known that fatty, membranous materials (such as lipids) can organise into a variety of shapes (phases), including stacked sheets (lamellae), tubes, or cubes, to name a few. Controlling these phases is essential to proper function. This new information will allow the researchers to identify critical components involved in neurodegenerative diseases such as multiple sclerosis (MS).
MS is an autoimmune disease resulting in the destruction of myelin, a fatty substance that insulates nerves and increases the speed at which signals travel between nerve cells. MS affects more than 2.3 million people worldwide and has no cure. By investigating the microscopic structure of myelin membrane under various conditions, the researchers were able to identify the critical conditions that alter the myelin structure. Their results showed a phase transition from a healthy stack of lamellas to a diseased inverted hexagonal phase as a result of the altered lipid stoichiometry, myelin basic protein content, and environmental conditions such as salinity and temperature. Myelin lipid composition, and the physiological environmental conditions, are critical for myelin to function properly, and the results demonstrate that these conditions should be contemplated as alternative routes for MS early detection.
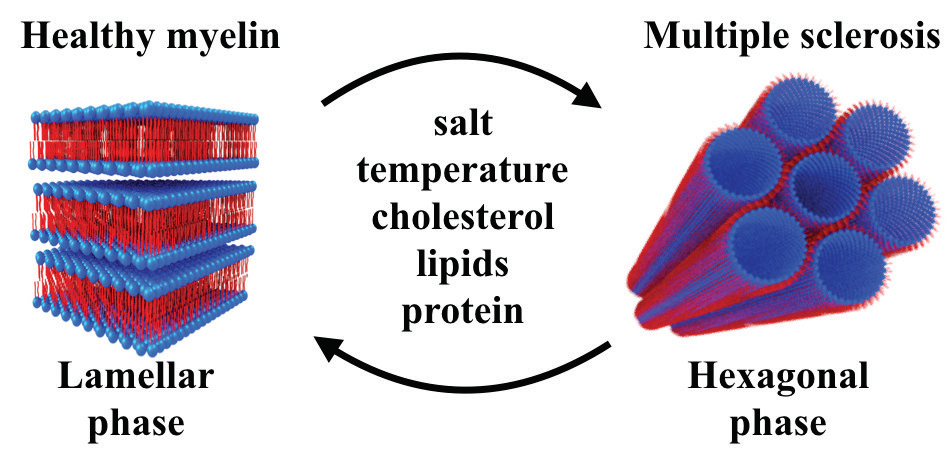
The myelin sheath, a repeating lipid-protein multilamellar structure, acts as an electrical insulator by forming a capacitor surrounding the axon, insuring fast nerve conduction1. The myelin membrane is composed of multiple lipid types, and the myelin basic protein (MBP). MBP facilitates binding of the myelin sheath to the underlying cytoplasmic membrane presumably through a charge-charge interaction where the positively-charged protein binds to the negatively-charged membranes acting as an intermolecular adhesion glue2,3.
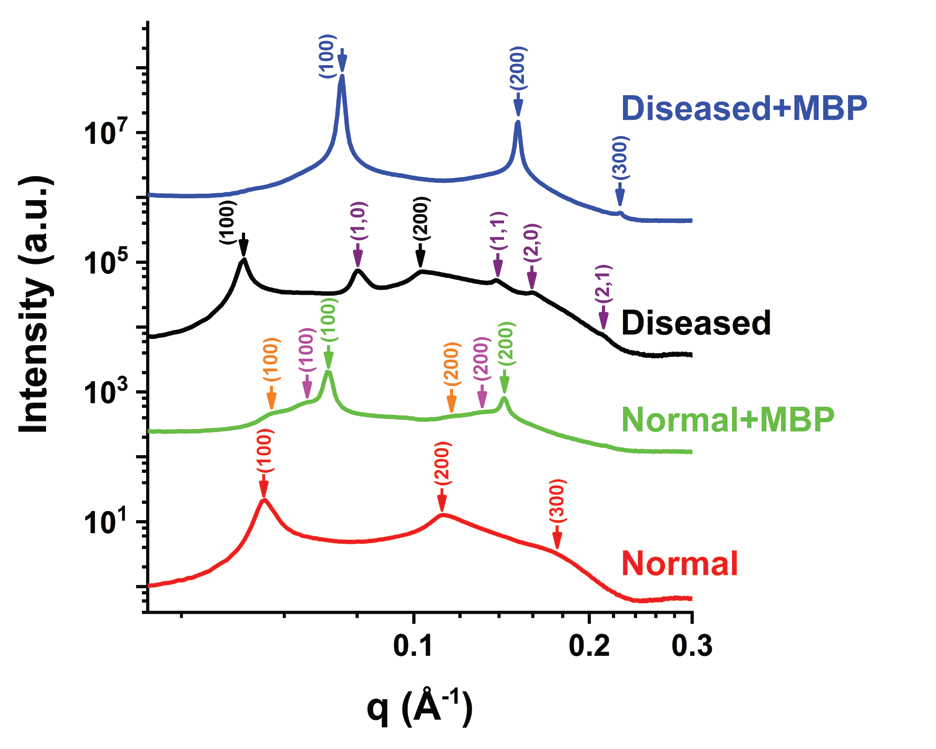
In vivo, the environmental conditions are highly regulated and controlled. Potassium and sodium are essential for proper action potential along the myelinated axons. Calcium and zinc ions have the same oxidation state, yet they have different biological functions in the body, such as signal transduction pathways, and the induction of protein-protein interactions. Due to the fact that the myelin membrane is close to a structural phase transition, we addressed the effects of altered environmental conditions on the cytoplasmic myelin membrane structures5.
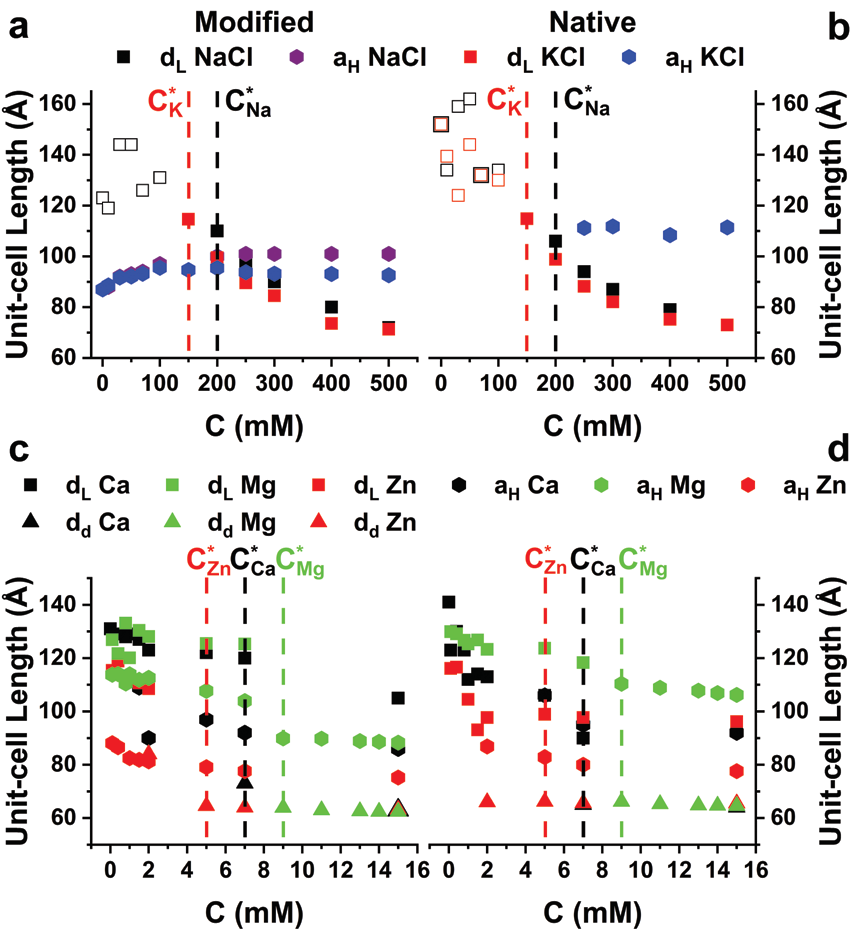
The divalent ions results are dramatically different. Here, we find that the structures are much more ion-specific (Fig. 3c-d). For example, the inverted hexagonal phases differ in the unit-cell length sizes at saturation with aH (Mg2+) > aH (Ca2+) > aH (Zn2+). Moreover, above a critical concentration, C*(Zn2+) = 5 mM < C*(Ca2+) = 7 mM < C*(Mg2+) = 9 mM, we find a coexisting dense lamellar phase (dd). This new phase has the same unit-cell length for all ion types and lipid compositions (dd ∼ 64 Å). Moreover, above C*, we find no changes in the hexagonal unit-cell lengths (Fig. 3, hexagonal symbols). Surprisingly, here we find that even the normal lipid composition exhibits coexisting of inverted hexagonal phases, similar to the diseased state.
In order to study myelin vulnerability, we measured its structure in the context of membrane self-assembly by SAXS. From a physical perspective, the competing forces between the lipids determine the nanoscopic structure and the macroscopic mesophase, discussed in detail in ref4,5. In these studies, we identified several factors that tend to destabilise the lamellar phase, and induce the formation of the inverted hexagonal phase. Minor alterations of the environmental conditions can drive structural instabilities and the formation of the HII phase. Lower salinity and low temperature are favorable for healthy lamellar phase up to about physiological condition.
In summary, even healthy myelin lipid composition is on the verge of a structural phase transition. The phase transition can be linked as a possible trigger for the outbreak of MS. In the process of phase transition from lamellar into the HII phase, the membrane undergoes large undulations typical to the increase in local curvatures5. These undulations can induce spontaneous pores, resulting in the vulnerability of the membrane to an attack by the immune system. Therefore, changes in lipid composition, depletion of MBP, or local environmental modification near the myelin, whether by ion type or ion concentration, can result in pathological phase transition that characterises the diseased state.
References:

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Copyright © Diamond Light Source. Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.