 | ||
A structure for the enigmatic hepatitis A virus |
Beamlines I03/I24 Scientific Highlight
Hepatitis A virus (HAV) infects humans, targeting the liver. Despite a successful vaccine there are still around 1.4 million cases annually and a significant number of fatalities1. Unlike hepatitis B and C, it is classified as a pico-RNA-virus having an un-enveloped 300 Å diameter outer protein shell (capsid), which encloses a single-stranded RNA genome. The two ends of the genome are not translated, but the vast majority is read-off to a single polypeptide, cleaved initially into three poly-proteins (P1-P3). P1 is subsequently cleaved to yield the structural proteins (VP0, 1 and 3), which self-assemble into a spherical capsid with icosahedral (532) symmetry. VP0 is finally cleaved by a poorly understood mechanism to yield VP2 and an N-terminal fragment VP4.
The broad structure of picornavirus capsids is well characterised: VP1-3 assume an eight-stranded anti-parallel ‘β-barrel’ fold with the N-termini at the inner surface and the C-termini external, VP4 is internal and partly ‘invisible’ to structural analyses since it may be flexible or contacting the RNA which does not adhere to strict icosahedral symmetry. Five copies of VP1 associate around the five-fold-axes and three copies of VP2 and VP3 alternate around the three-foldaxes. VP2 abuts at the two-fold-axes. The external surface mediates interactions with the host organism: with cell surface receptors to gain entry into cells and with the cellular and molecular components by which the host’s immune system protects against pathogens. The surface loops have little structural constraint thus tolerating insertions, deletions and mutations that enable the virus to evade immune surveillance. Conversely viruses of one major genus – enterovirus – have a depression (canyon) around the five-fold-vertices, which frequently harbours the site of receptor interaction, near which is an opening into a hydrophobic pocket in the VP1 β-barrel in which a lipid (pocket-factor) binds. Receptor binding dislodges the lipid leading to the collapse of the pocket and concomitantly with uptake into cellular vesicles, structural re-arrangements occur opening up channels both on and adjacent to the two-fold-axes such that the internal hydrophobic protein segments (VP4 and the N-terminus of VP1) are discharged to engage with and form channels in the vesicle membrane, somehow triggering the release of RNA which passes into the cytoplasm for replication and the production of progeny virus2. A similar mechanism may apply for other genera of the Picornaviridae but the details are unclear.
This work on HAV provides the first structural information on the hepatovirus genus. HAV has radically different physico-chemical properties to other picornaviruses3 and has been relatively little studied. Recently it was shown that the virus exists in an enveloped form (garnering membrane from the host) whilst outside cells, potentially shielding itself from immune surveillance4. HAV is remarkably stable both genetically and physically being able to withstand temperatures up to 80 °C and pH as low as two, making disinfection tricky. HAV evaded structural analysis for many years since it is difficult to grow – unlike other picornaviruses it does not shut down host protein synthesis and the genome contains many rare codons – and its ‘stickiness’ made it very difficult to crystallise. HAV appeared to have a very short (23 amino acids), non-myristoylated VP4 although it had not been conclusively identified. Unlike other picornaviruses the capsid assembles a fragment of protein (VPX) that would be expected to belong to P2 (protein 2A in the other viruses) still attached as an extension to VP1. VPX is likely to mediate the association with membrane envelope, and is later cleaved off by host proteases to yield the mature, unenveloped capsid.
With inactivated vaccine virus material (formaldehyde treated) from the Sinovac company, thin, plate-like crystals were grown for both the mature and natural empty virus (capsids which did not package RNA). The group tried to collect data at room temperature by in situ diffraction i.e. putting the crystallisation plate in the beam with the crystals in situ in their mother liquor, however, the MPD in the mother liquor caused the crystal to ‘jump’ out of the beam when exposed, rendering data collection impossible. Data were therefore collected from cryo-cooled crystals on beamlines I03 and I24. The higher mosaic spread was ameliorated by the low beam divergence on I03. Typically 50 useful images could be collected from one position of a crystal and of the 200 crystals tested some 17 empty and 32 mature virus yielded data, and structures were solved at 3.5 Å and 3.0 Å respectively.
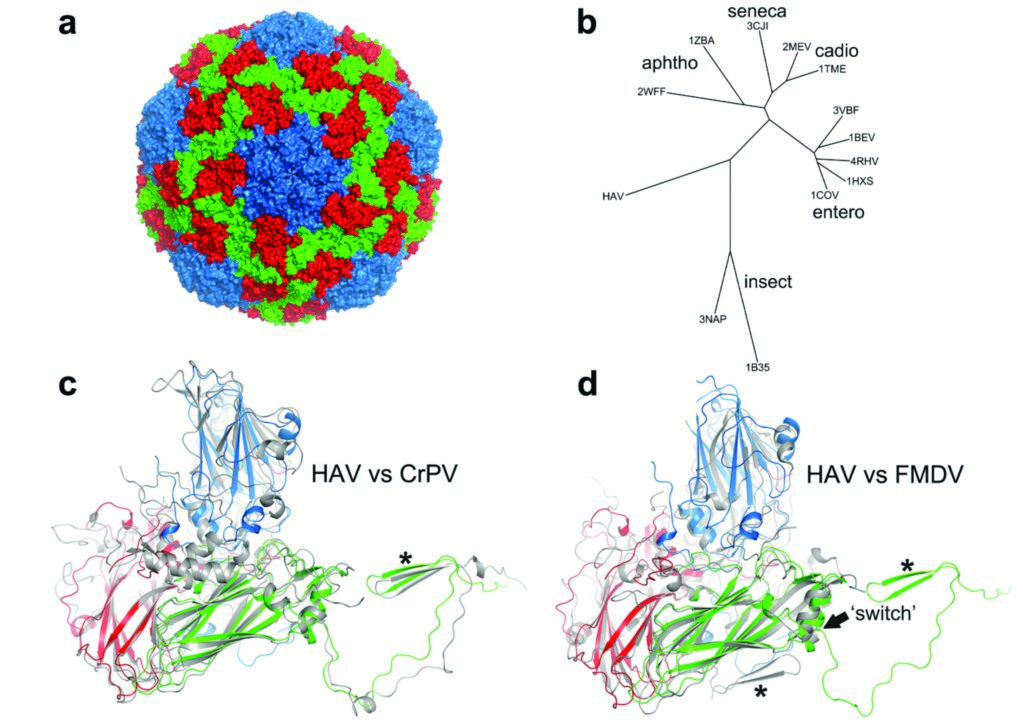
The HAV particle resembles other picornaviruses, most closely the aphthoviruses, although HAV is more facetted than the rather smooth spherical aphthoviruses. There are no major depressions suggestive of receptor attachment sites (Fig. 1). Similarly, HAV VP1 does not possess the hydrophobic pocket found in enteroviruses (the volume is largely filled with hydrophobic side chains). Most significantly there is a change in the structure at the two-fold axis compared to other mammalian picornaviruses, a domain swap for the N-terminus of VP2 whereby the same β-hairpin structure is formed at the two-fold symmetric position. This same structure is seen in insect picorna-like viruses5. These factors strongly suggest that HAV uncoats via a novel mechanism. The mature and natural empty capsids are almost identical externally but internally, the empty capsid has more disordered protein corresponding to regions of VP0 and neighbouring stretches of the N-terminus of VP1. Whilst VP4 is seen on an SDSPAGE of the mature capsids, it is not visible in the structure. Although it has some hydrophobic character it is hard to see how this shortened, non-myristoylated version of VP4 could fulfill the same role as in the enteroviruses (engaging with and forming a pore in the endosome membrane). The surface of the virus is remarkably negatively charged and the fringes of the pentameric assemblies, which show some positive charges, are decorated with a string of sulphate ions derived from the crystallisation media. Reported neutralising epitopes have been mapped onto the capsid and we have evidence implicating further residues in VP3 and VP2.
Source publication:
Wang, X., Ren, J., Gao, Q., Hu, Z., Sun, Y., Li, X., Rowlands, D. J., Yin, W., Wang, J., Stuart, D. I., Rao, Z. & Fry, E. E. Hepatitis A virus and the origins of picornaviruses. Nature 517, 85-U214, doi:10.1038/nature13806 (2015).
References:
1. WHO. Hepatitis A. Fact Sheet No. 328 http://www.who.int/mediacentre/ factsheets/fs328/en/ (2014).
2. Wang, X. et al. A sensor-adaptor mechanism for enterovirus uncoating from structures of EV71. Nature Structural & Molecular Biology 19, 424-429, doi:10.1038/nsmb.2255 (2012).
3. Martin, A. & Lemon, S. M. Hepatitis A virus: From discovery to vaccines. Hepatology 43, S164-S172, doi:10.1002/hep.21052 (2006).
4. Feng, Z. et al. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 496, 367-71, doi:10.1038/nature12029 (2013).
5. Tate, J. et al. The crystal structure of cricket paralysis virus: the first view of a new virus family. Nature Structural & Molecular Biology 6, 765-774, doi:10.1038/11543 (1999).
Funding acknowledgements:
This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences, grant no. XDB08020200, the Ministry of Science and Technology 973 Project (grant no. 2014CB542800), the National Major Project of Infectious Disease (2012ZX10004701) and National Science Foundation grant 81330036. D.I.S. and E.E.F. are supported by the UK Medical Research Council (grant G1000099) and J.R. by the Wellcome Trust. This work is a contribution from the Instruct Centre, Oxford. Administrative support was provided by the Wellcome Trust (075491/Z/04).
Corresponding author:
Dr Elizabeth Fry, Oxford University, [email protected]
