
HIV is a retrovirus that needs to copy its RNA content into DNA within a host cell. How it avoids being detected and destroyed by our immune system during this process is currently unknown, and fully understanding this mechanism is paramount to developing an effective HIV vaccine.
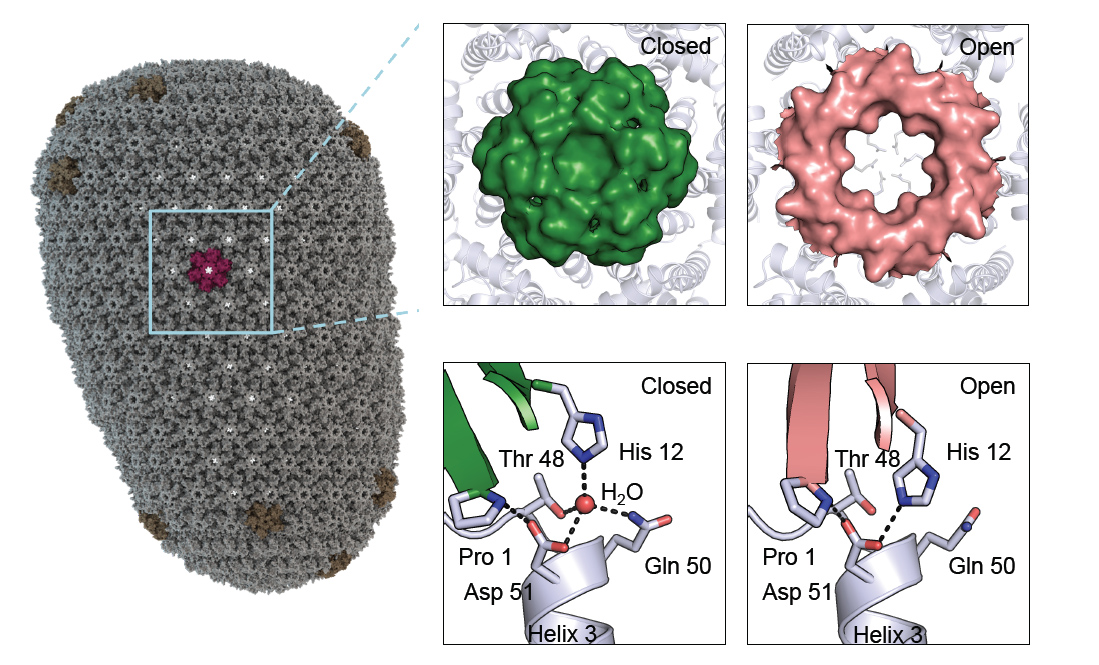
Human immunodeficiency virus (HIV-1) is the etiological agent behind AIDS. According to WHO statistics, HIV-1 affects approximately 40 million people worldwide, kills more than 1 million every year and newly infects over 2 million. While antiviral agents are available, if costly, there is no vaccine and a rise in resistant strains, particularly in sub-saharan Africa where ~40% of adults are on antiviral therapy.
Despite repeated and ongoing zoonotic transmissions into human populations, it is a single strain, HIV-1 M, that is responsible for the current pandemic. Intense research effort has therefore been made to understand what makes this strain so deadly to humans. HIV-1 is a retrovirus, meaning that it must reverse transcribe its RNA genome into DNA before it can begin replicating. This process of reverse transcription happens after the viral capsid enters the cell, but the cytosol is home to potent nucleases and immune sensors that detect and destroy foreign DNA1. Furthermore, while the viral reverse transcriptase enzyme and RNA genome are located inside the capsid, the nucleotides that fuel DNA synthesis are located outside. This creates a paradox long unsolved in HIV biology; if the virus uncoats it gains access to nucleotides but becomes exposed to the cytosol, if it doesn’t it remains protected but cannot reverse transcribe. Although many capsid structures have been solved over the years, by Nuclear Magnetic Resonance (NMR), crystallography and cryo-electron microscopy (cryo-EM), they have provided no clues to this puzzle.
The first clue came when HIV-1 capsid hexamers were trapped in several alternative conformations. Data collected at Diamond’s I02 beamline revealed the existence of a dynamic pore at the centre of the 6-fold axis (Fig. 1). The pore is formed by an N-terminal β-hairpin which hinges around a hydrogen bond between a histidine (H12) and an aspartate (D51). In one conformation, which can be trapped under basic pH conditions, H12 is deprotonated and this interaction is water-mediated, forcing the hairpin into the centre of the hexamer and closing the pore. Under neutral pH conditions where H12 is protonated, a direct hydrogen bond is made with D51, pulling the hairpin back and opening the pore. Crucially, all previous hexamer structures had been solved under basic deprotonated conditions, hiding the pores existence. The significance of this discovery is only fully apparent when the hexamer is used to build a model of an intact virion: the HIV-1 capsid is a dynamic, highly porous structure.
Further clues emerged when the hexamers were analysed in more detail. A cross section through the protein revealed a hidden chamber with a volume of > 3,000 Å, approximately 10 times that of a deoxynucleotide triphosphate (dNTP) (Fig. 2). At one end, the β-hairpin acts as a gatekeeper, controlling access to the chamber by anything outside. At the other end, a ring of arginine residues (R18) lines the exit into the capsid interior. Hypothesising that this might be the means by which dNTPs are recruited into the capsid to fuel DNA synthesis, evidence for direct interaction was sought. Remarkably, HIV-1 capsid was found to not only bind dNTPs but to do so with an affinity of ~30 nM. While such high affinity is the hallmark of a specific interaction, tight binding is not necessarily a favourable feature in an import mechanism. Kinetic analysis provided the answer; while the affinity is high the off-rate is fast. Nucleotides are bound with a half-life of only 58 ms. The apparent high affinity is a result of the efficiency with which the capsid attracts dNTPs. In fact, the on-rate for this step is diffusion limited, meaning that the HIV-1 capsid has evolved to recruit nucleotides as fast as physical laws allow. Re-solving the hexamer in the presence of deoxyadenosine triphosphate (dATP) confirmed the theory; the positively charged ring of arginine residues recruits negatively charged triphosphates via electrostatic steerage (Fig. 2).
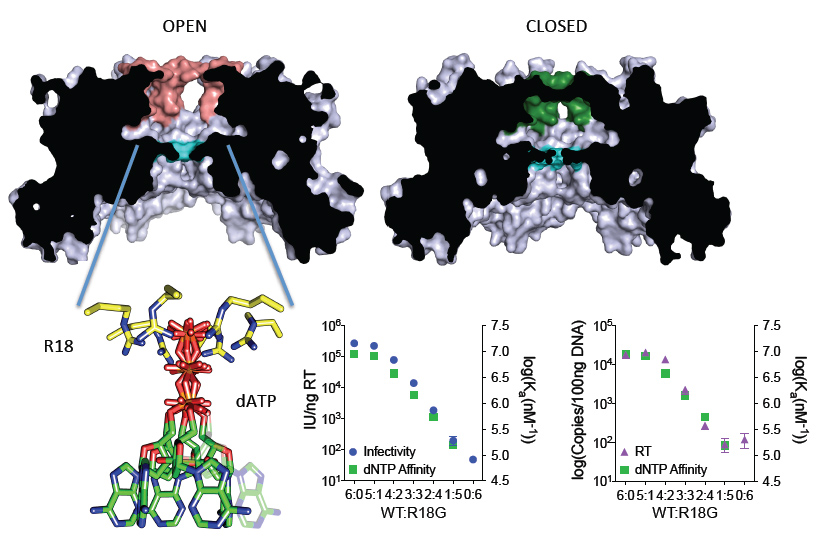
Figure 2: Capsid nucleotide channels are essential for HIV infection. The HIV capsid is comprised of hexameric units. At the centre of each hexamer is a hidden chamber through which nucleotides are imported. Nucleotides are attracted by a ring of arginine residues, which coordinate the negatively charged phosphates. When the arginine residues are removed, HIV cannot synthesise DNA and becomes non-infectious.
While compelling, structural data alone cannot prove biological significance. To demonstrate that dNTP import by the capsid is an essential step during HIV-1 infection binding was correlated with infectivity and reverse transcription. First, chimeric hexamers were created by mixing different ratios of wild-type and R18G mutant capsids, disassembling them into monomers then allowing them to re-oligomerize. This generated a series of chimeras in which R18 was incrementally replaced with glycine. Repeating the dNTP binding experiments with these chimeras revealed that binding was proportional to the number of arginine residues in the pore. Meanwhile, a matching set of chimeric HIV-1 viruses were produced by varying the expression of wild-type and R18G capsid in virus producing cells. Strikingly, sequential removal of R18 resulted in a dose-dependent decrease in both reverse transcription and infection. Combining these datasets produced a conclusive correlation; HIV-1 infectivity is dependent upon efficient dNTP recruitment by the R18 capsid pore (Fig. 2). The dependence of HIV-1 on dNTP recruitment reveals the capsid pore as a potential new drug target for antiretrovirals. Using the hexamer structures to rationally select drug candidates, a series of carboxybenzene compounds were identified that could compete with dNTPs and prevent reverse transcription in vitro. Solving a complex between one such compound, hexacarboxybenzene, and capsid hexamer revealed that the interaction mimics a bound dNTP but engages all six arginines simultaneously (Fig. 3). Crucially, interaction is only through these arginines, meaning that evolving drug resistance would be exceptionally challenging for the virus.
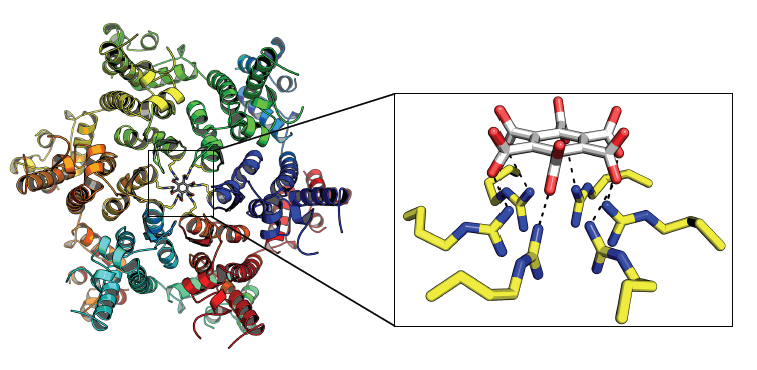
Figure 3: HIV capsid pores can be blocked with small molecule drugs to prevent nucleotide import and viral DNA synthesis.
Structural data collected at Diamond has finally allowed us to unravel a mystery at the centre of HIV-1 biology. It has revealed that the capsid, once thought to be a static and impervious shell, is actually highly dynamic and selectively permeable. Pores at the centre of each hexameric subunit recruit dNTPs into the capsid interior whilst keeping out nucleases and immune sensors that would inhibit infection. This mechanism helps explain how HIV- 1 infects cells whilst evading the host immune response, with devastating consequences for the global population.
References:
1. Rasaiyaah, J. et al. HIV-1 evades innate immune recognition through specific cofactor recruitment. Nature 503, 402- 405, doi:10.1038/nature12769 (2013).
Funding acknowledgement:
This work was supported by the Medical Research Council (UK; U105181010)
Corresponding author:
Dr Leo James, MRC Laboratory of Molecular Biology, lcj@mrc-lmb.cam.ac.uk
Related publication:
Jacques DA, McEwan WA, Hilditch L, Pice AJ, Towers GJ, James LC. HIV-1 uses dynamic capsid pores to import
nucleotides and fuel encapsidated DNA synthesis. Nature 536, 349-353, doi:10.1038/nature19098 (2016).
Publication keywords:
HIV; Capsid; Innate immunity; Viral infection

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Copyright © 2022 Diamond Light Source
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.

 Science
Science