
The anaphase promoting complex or cyclosome (APC/C) is a multi-subunit cullin-RING E3 ubiquitin ligase that functions to regulate progression through the mitotic phase of the cell cycle and controls entry into S phase 1. APC/C-mediated coordination of cell cycle progression is achieved through the temporal regulation of APC/C activity and substrate specificity. The core APC/C is assembled from 13 different proteins, mostly highly conserved and essential for function, generating a macromolecular machine exceeding 1.2 MDa in mass. The catalytic centre of the APC/C is formed from the cullin and RING subunits Apc2 and Apc11, analogous to Cul1 and Rbx1, respectively, of the SCF cullin RING ligase. In combination, Apc2 and Apc11 are competent to catalyse protein ubiquitylation, although lack substrate selectivity. Single particle electron microscopy studies of yeast and vertebrate APC/C, revealing the overall shape, architecture and dimensions of the molecule have been supplemented by crystallographic studies of a few individual APC/C subunits. A particularly striking feature of APC/C subunits is that many incorporate multiple sequence repeats. The most prevalent structural motif is a degenerative 34-amino acid unit termed the tetratricopeptide repeat (TPR). Four conserved APC/C subunits (Cdc16, Cdc23, Cdc27 and Apc5), together with Apc7 unique to vertebrate APC/C, are almost exclusively composed of multiple TPR motifs, usually arranged in contiguous arrays. In contrast, Apc1, the largest APC/C subunit, consists of nine tandem repeats of a 30 – 40 residue proteasome-cyclosome motif, whereas the N-terminus of Apc2 comprises three consecutive ~130 residue cullin repeats. Multiple TPR arrays, originally discovered within proteins subsequently identified as components of the APC/C, and now recognised as ubiquitous to all domains of life, form superhelical structures that function to mediate protein–protein interactions, particularly the assembly of multiprotein complexes.
Our overall aims are to determine a high-resolution structure of the APC/C to address questions of its assembly and molecular basis for substrate specificity and catalysis, and how the APC/C is regulated at different phases of the cell cycle.
To understand the structure and function of the APC/C’s TPR subunits, we crystallised an S. pombe Cdc16-Cdc26 subcomplex. The structure was determined by means of SAD phasing on a gold derivative using data collected at beam-line I02, Diamond Light Source 2. Cdc16 homo-dimerises through its N-terminal seven TPR motifs, by means of an inter-locking TPR–TPR homotypic interaction essentially identical to the N-terminal dimerisation domain recently reported for Cdc273 (Fig. 1). Cdc16’s appearance is dominated by the rod-like structure of each Cdc16 protomer. Cdc16 is composed virtually entirely of 14 contiguous TPR motifs (including two non-canonical TPR motifs). Each TPR motif consists of a pair of anti-parallel a-helices, termed A and B, rotated relative to one another by ~24°. Successive A and B a-helices generate a right-handed TPR superhelix with A-helices lining the inner concave groove and B-helices forming the outer convex surface. In Cdc16, the 14 TPR motifs together with a C-terminal capping a-helix (a29) generate 29 antiparallel a-helices that adopt a linear superhelix measuring some 110 Å in length and 60 Å in diameter. Cdc16 subunits homo-dimerise through their Cdc16Nterm domains to generate a shallow ‘V’-shaped molecule with a maximum dimension of some 155 Å The more globular Cdc16Nterm dimerisation module forms the apex of the ‘V’ with the narrower C-terminal TPR superhelices of Cdc16Cterm projecting away from the dimer interface. Cdc16Cterm contributes the majority of Cdc16 interactions to Cdc26. The Cdc16 dimer interface is structurally related to the N-terminal dimerisation domain of Cdc273 , demonstrating that both Cdc16 and Cdc27 form homo-dimers through a conserved mechanism (Fig. 2).
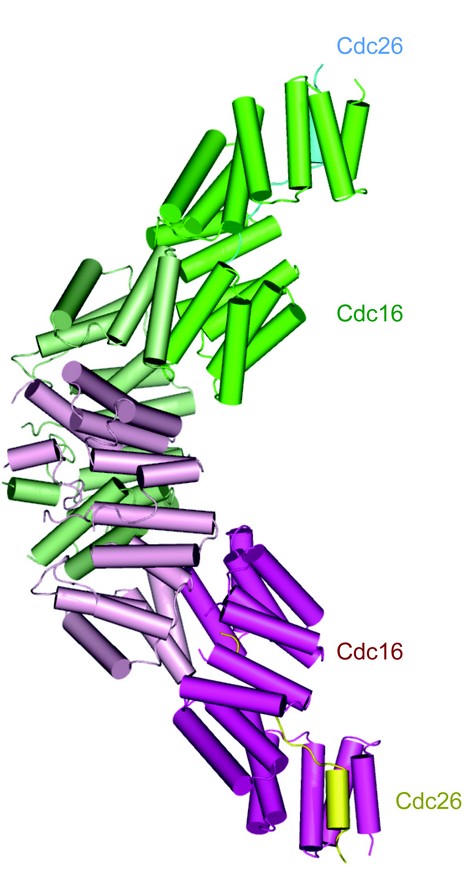
Figure 1: View of the Cdc16-Cdc26 heterotetramer2
The N-terminal 13 residues of Cdc26 form an extended conformation running parallel with, and lining the inner groove of the Cdc16 TPR superhelix contacting, primarily, although not exclusively, A-helices of Cdc16Cterm (specifically TPR7 – TPR14). Residues 14 - 21 of Cdc26 fold into an a-helix that packs parallel with a29 of Cdc16, augmenting the array of Cdc16 a-helices. Electron density for the acetyl modification of Met1 of Cdc26 is clearly visible in the 2Fo_Fc map. The N-terminal acetylated-Met residue is located within an enclosed chamber formed from the inner groove of the Cdc16 TPR superhelix the N-terminus of Cdc26 is completely surrounded by its interacting Cdc16 subunit. The upper extent of the chamber is capped by the N-terminus of the opposing Cdc16 subunit, although Cdc26 does not participate at the dimer interface. Thus, the acetyl group of the N-terminal Met of Cdc26 is completely inaccessible to bulk solvent.
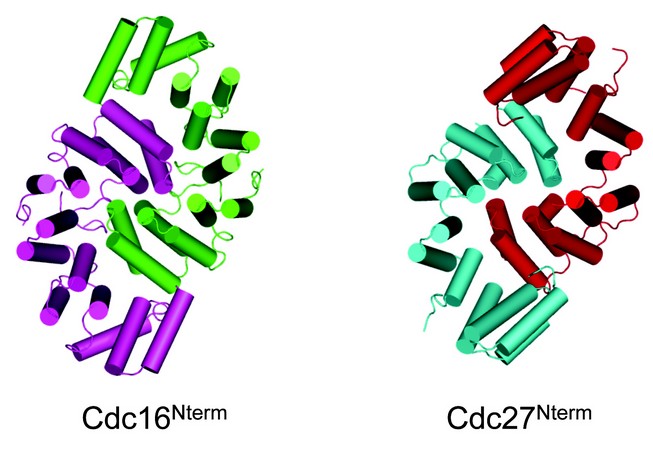
Figure 2: The N-terminal dimerisation domains of Cdc26 and Cdc27 are structurally equivalent 2,3.
The crystal structure of the N-terminal domain of Encephalitozoon cuniculi Cdc27 (Cdc27Nterm) was determined using SAD SeMet data collected at beam line I02, Diamond Light Source 3. E. cuniculi Cdc27Nterm comprises 14 a-helices and forms a homo-dimer adopting a shape resembling a shallow elongated bowl measuring 75 Å x 50 Å x 30 Å. The core architecture of each subunit is formed from 13 a helices organised into an antiparallel arrangement, closely resembling a TPR fold.
We used the crystal structure coordinates of Cdc16-Cdc26 and Cdc27 and other previously determined crystal structures of APC/C subunits and homology-derived coordinates 1 to interpret a 10 Å resolution cryo-electron microscopy map of the APC/C in complex with its co-activator Cdh1 and a D-box-substrate.4,5 This provided a precise definition of the organization and structure of all essential APC/C subunits, resulting in a pseudo-atomic model for 70% of the APC/C (Fig. 3). To determine the position of individual APC/C subunits we developed an overexpression system for the APC/C that reconstitutes all 13 APC/C proteins generating a functional E3 ligase. This system allowed us to determine the APC/C mass and subunit stoichiometry accurately and delineate the molecular boundaries of most APC/C subunits within the EM-derived molecular envelope. A lattice-like appearance of the APC/C is generated by multiple repeat motifs of most APC/C subunits. Three conserved tetratricopeptide repeat (TPR) subunits (Cdc16, Cdc23 and Cdc27) share related superhelical homo-dimeric architectures that assemble to generate a quasi-symmetrical structure. Our structure explains how this TPR sub-complex, together with additional scaffolding subunits (Apc1, Apc4 and Apc5), coordinate the juxtaposition of the catalytic and substrate recognition module (Apc2, Apc11 and Apc10), and TPR-phosphorylation sites, relative to co-activator, regulatory proteins and substrates.
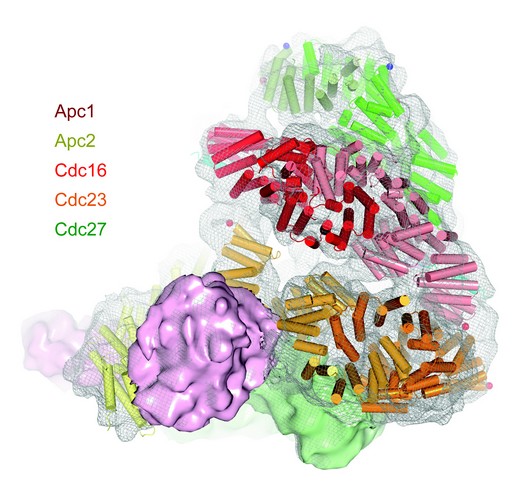
Figure 3: Pseudo-atomic model of the APC/C. Crystal coordinates and homology-derived coordinates were fitted to a 10 Å cryo-EM map of the APC/C 4,5.
In the future we are pursuing efforts to crystallise the remaining APC/C subunits and sub-complexes and the entire APC/C assembly.
Zhang Z, Kulkarni K, Hanrahan SJ, Thompson AJ, Barford D.The APC/C subunit Cdc16/Cut9 is a contiguous tetratricopeptide repeat superhelix with a homo-dimer interface similar to Cdc27. EMBO J. 29(21) 3733-44 (2010)
References

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Copyright © 2022 Diamond Light Source
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.

 Science
Science