
Dr Geoff Sutton, University of Oxford
Virus infections in a number of insect families generally culminate in the occlusion of between one and several thousand virus particles within large proteinaceous crystalline occlusion bodies or polyhedra. Polyhedra are generally less than 7 μm in size, serve to protect the viruses from harsh environmental conditions and are made from a protein, polyhedrin, encoded by the viruses. Despite their small size, we solved the structure of polyhedra produced by the dsDNA baculovirus, Autographa californica multiple nucleopolyhedrosis virus (AcMNPV), with data collected on the Diamond microfocus beamline I24 to 1.8 Å resolution.
The structure reveals an intricate packing which forms a highly symmetrical, covalently cross-braced lattice. The polyhedrin subunits form tightly packed trimers which are then linked as dodecamers by disulphide bonds. The dodecamers interlock to form the robust crystal. A flexible adaptor enables this supra-molecular assembly to specifically entrap massive baculoviruses. Inter-subunit chemical switches modulate the controlled release of virus particles in the unusual high pH environment of the target insect’s gut.
Surprisingly, the structure of the AcMNPV polyhedrin subunits is more similar to the jelly roll fold of picornavirus coat proteins than to the polyhedrin of a dsRNA virus cytoplasmic polyhedrosis virus (BmCPV1). It is, therefore, remarkable that despite sharing little amino acid sequence identity, both AcMNPV and BmCPV1 polyhedra possess identical crystal lattices and crystal symmetry, however the structures are, at best, distantly related. The crystalline arrangement must be particularly well suited to the functional requirements of the polyhedra and has been either preserved or re-selected during evolution. The use of flexible adaptors to generate a powerful system for packaging irregular particles is characteristic of the AcMNPV polyhedrin and may provide a vehicle to sequester a wide range of objects such as biological nano-particles.
Insect viruses have developed methods to protect their virions when outside the host organism, a striking example being the sequestration within robust crystalline particles. These occlusion bodies (or polyhedra) vary in size (typically 0.1-5 µm), often have characteristic shapes and well ordered lattices, and are built primarily by a single protein encoded by specific groups of viruses including baculoviruses, cytoplasmic polyhedrosis viruses (CPVs) and entomopoxviruses. They act as protective packages allowing infectious virions to survive for long periods in harsh environments, providing a delivery system between hosts by oral-faecal routes and resisting solubilisation until exposed to the alkaline insect midgut. The fact that these diverse groups of insect viruses all use occlusion as a method to package their virions suggests that there may be a powerful selective advantage for their survival within insects.
Baculoviruses are a family of large dsDNA viruses which replicate and assemble within the host nucleus. The baculovirus polyhedra protein, polyhedrin, is one of the most conserved proteins in the virus. Naturally occurring aminoacid substitutions, many of them single point mutations, produce a variety of phenotypic changes to the ranging from large, cuboid polyhedra, which occlude no or few virions, to overall changes in polyhedra shape.
We have determined, from in vivo grown crystals, the structure of both virus-containing wild-type and a virus-empty mutant (G25D) polyhedra from the baculovirus Autographa californica multiple nucleopolyhedrosis virus (AcMNPV). The crystals were derived from cultured insect cells with the wild-type crystals being usually <5 µm in maximum linear dimension while many G25D crystals attained 5-10 µm. Despite their small size, we were able to determine the high resolution (1.8 Å) structure for the mutant polyhedra using conventional seleno-methionine labelling to solve the phase problem by taking data on the tuneable microfocus beamline I24. Due to the small size of the crystals and their low solvent content (nominally 21%), structure determination was challenging; however, the final structure is reliable (Rwork=0.164, Rfree=0.217 from 36.3 to 1.8 Å resolution). Two major disordered regions are present where the residues could not be positioned; D1 from 32-48 and D2 from 174- 186.
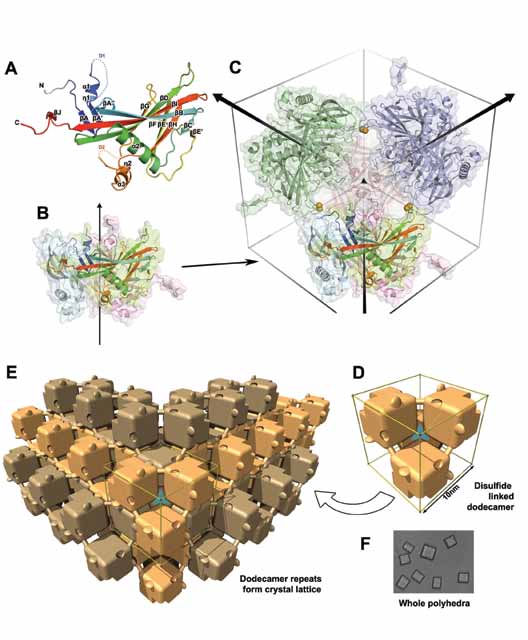 |
| Figure 1 (A) Cartoon representation of the structure of AcMNPV polyhedrin coloured from blue at the N-terminus to red at the C-terminus. Disordered regions D1 (residues 32-49) and D2 (residues 174-186) are shown as dots. (B) Cartoon and surface representation of polyhedrin trimer aligned to the orientation in (C). Subunits are coloured by chain with one in the trimer coloured as in (A). (C) Dodecameric unit of four trimers linked by disulphide bonds (represented as orange spheres). The unit cell and threefold axes are drawn to facilitate observation. (D) Schematic of disulphide linked dodecamer, orientated as in (C). Each trimer is represented as a simplified cubic block (with one corner absent), with the C-terminal hooks and pockets present. The disulphide linking the trimers is represented as a dowel. (E) The crystal lattice is built up from repeats of the dodecameric unit, alternately coloured beech and brown to aid viewing. (F) Light microscopy image of G25D mutant AcMNPV polyhedra. |
AcMNPV polyhedra have I23 symmetry with one polyhedrin subunit in the asymmetric unit, so that 24 copies of the 28.6 kDa (245 amino acids) molecule are tightly packed in the unit cell (a,b,c=102 Å). The polyhedrin subunit can be split up into 3 parts: N-terminal head, a β-barrel body (which comprises most of the molecule) and a C-terminal tail (Figure 1A). The β-barrel body is reminiscent of that found in the major capsid proteins of picorna-like viruses. The subunits form tightly packed trimers, with the disordered portions of the molecule grouping at one corner of the essentially cubeshaped assembly (Figure 1B). The jelly-roll β-barrels of each subunit are packed orthogonal to each other and define the basic size of the crystal building block. The N-terminal head contributes to the clamping sheet fundamental to the trimer assembly, part of a robust trimer interface comprising a mixture of hydrophobic interactions, salt bridges and hydrogen bonds, which buries ~25% of the surface area of each polyhedrin subunit. The C-terminal hooks protrude at right angles from three edges of the trimer. Eight of the cubelike trimers are neatly arranged to fill the crystal unit cell. There are in fact two nested sets of four trimers, one arrayed at the corners of the cell and the other at the centre; four trimers point inwards towards the centre of the cell, whereas the other four point out towards the corners. The closest interactions between the trimers of the same polarity come at the centre of the face of the unit cell. This contact is not extensive (~800 Å2) and would not normally be sufficient to achieve a strong interaction; however, four salt bridges between highly conserved glutamate, arginine and asparagines side chains and a covalent disulphide bond between 2 symmetry-related copies of Cys132 tether this extended dimer. As Cys132 is the only cysteine fully conserved across the baculoviruses it seems likely that this disulphide stabilization is biologically relevant. These interactions link four trimers of the same polarity in a tetrahedral arrangement to form a dodecameric cage (Figure 1C, D). The assembly of the crystal is completed by clipping together dodecamers (Figure 1E). In this lattice the disulphide bonds provide cross-braces, whereas stability in the direction of the unit cell edges is provided by a very extensive interface (~10,000 Å2). This includes the protruding C-terminal hooks which engage adjacent dodecamers through a series of conserved residues. By light microscopy, the AcMNPV polyhedra appear as cubes (Figure 1F).
Overall the structure explains certain key physical properties of the polyhedra, revealing a robust isotropic structure with virus engagement modules arrayed at high density to specifically attach the viruses to be occluded. This raises interesting possibilities for redirecting these interactions towards the encapsulation of other structures. The structure also explains the mechanism of lattice dissolution in the high pH environment of the insect mid-gut, by revealing a high concentration of tyrosine side-chains at molecular interfaces, forming a pH dependent molecular switch.
The structure also illuminates the evolutionary history of polyhedrins – although it raises almost as many questions as it answers. Firstly it is clear that the baculovirus protein is surprisingly different in structure to the equivalent protein from the type 1 cytoplasmic polyhedrosis virus. Although both of these proteins possess a jelly-roll, the baculovirus polyhedrin is in fact more similar to the major coat proteins of picornaviruses. We believe that this latter similarity is sufficiently strong to support the hypothesis that they evolved from a common ancestral protein, however at this stage we cannot tell if the two known polyhedrin structures are also related. With tricky questions of evolution such as this it will be interesting to see if the determination of additional structures will clarify the situation by identifying serial homologues.
Principal Publications and Authors
Ji X, Sutton G, Evans G, Axford D, Owen R, Stuart DI. (2010). How baculovirus polyhedra fit square pegs into round holes to robustly package viruses. EMBO J. 29, 505-14. Epub Dec 3, (2009).
Funding Acknowledgement
Medical Research Council UK and SPINE2-COMPLEXES LSHGCT-2006-031220.

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Copyright © 2022 Diamond Light Source
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.

 Science
Science