Prof David Barford, Institute of Cancer Research
Rho proteins are small GTPases that function to regulate cell motility, polarity, adhesion, cytoskeletal organisation, proliferation and apoptosis. Two distinct families of Rho guanine nucleotide exchange factors (GEFs), the Dbl homology (DH) and DOCK proteins, activate Rho GTPases through the exchange of GDP for GTP. In humans, DOCK proteins are organised into four subfamilies, characterised by their differing specificities for Rac and Cdc42, regulatory domains and associated subunits. All contain a GEF catalytic domain named the DHR2 domain of ~400 residues situated within the C-terminus. To understand the molecular mechanisms of guanine nucleotide exchange catalysed by DOCK proteins, we determined the mechanism of Cdc42 (a Rho family GTPase) activation by the novel GEFs of the DOCK family. The structure of the DOCK9DHR2-Cdc42 complex in the nucleotide-free and in the presence of GDP and GTP allowed us to identify a nucleotide sensor within the a10 helix of the DHR2 domain that contributes to release of GDP and then to discharge of the activated GTP-bound Cdc42.
Malignant tumour cells invade local tissues and migrate (metastasis) to sites separate from the primary tumour. Metastasis is the main factor accounting for cancer treatment failure and is responsible for 90% of cancer deaths. Thus, intense research efforts are directed at understanding the cellular and molecular basis for tumour cell motility and invasion. Eukaryotic cells undergo cell migration through dynamic changes in their actin-based cytoskeleton, in a process controlled by small GTPases of the Rho family, coupled to reversible protein phosphorylation. Rho GTPases inter-convert between inactive GDP-bound states and active GTP-bound states, through a process controlled by guanine nucleotide exchange factors (GEFs) that catalyse exchange of GDP for GTP at the nucleotide-binding site. Two distinct families of Rho GEFs, the Dbl homology (DH) and DOCK proteins, activate Rho GTPases through the exchange of GDP for GTP [1,4]. In humans, there are ~60 DH containing Rho GEFs and 11 DOCK family members. DOCK proteins have been implicated in the activation of Rac and Cdc42 in cell migration, morphogenesis and phagocytosis, and as important components of tumour cell movement and invasion [2.3]. Although the structural basis of nucleotide release catalysed by the DH family of Rho GEFs is now established, the molecular mechanisms of DOCK proteins was unknown. Moreover, the structural events that permit discharge of the activated GTPase when GTP binds to the nucleotide free GTPase in a GEF-GTPase complex had not been defined. To understand the molecular mechanisms of DOCK proteins, we determined the mechanism of Cdc42 (a Rho family GTPase) activation by the novel family of guanine nucleotide exchange factors of the DOCK family.
|
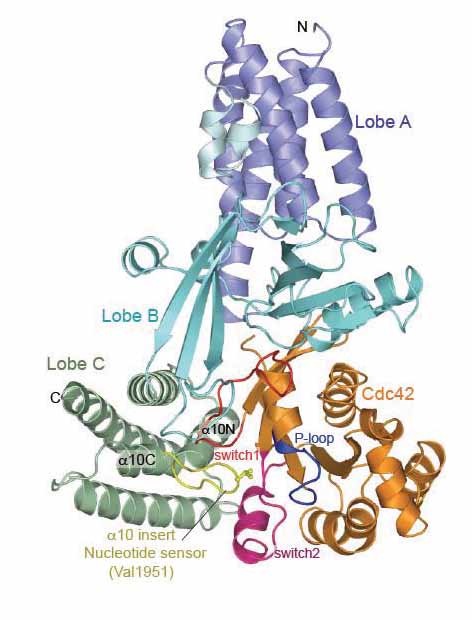
Figure 1. View of nucleotide-free DOCK9DHR2-Cdc42. | Specifically, we determined the structure of the catalytic domain of DOCK9 (DHR2 domain, DOCK9DHR2) in complex with Cdc42 and bound to GDP and GTP [5]. We found that the DHR2 domain, which has a different structure from DH GEF catalytic domains, is organised into three lobes of roughly equal size (lobes A, B and C), with the Cdc42-binding site and catalytic centre generated entirely from lobes B and C (Fig. 1). Lobe A is formed from an anti-parallel array of five a-helices that forms extensive contacts with lobe B to stabilise the DHR2 domain. Lobe B adopts an unusual architecture of two anti-parallel b-sheets disposed in a loosely packed orthogonal arrangement, whereas lobe C comprises a four-helix bundle. Helix a10 of lobe C, the most conserved region of DHR2 domains is interrupted by a seven-residue loop - the a10 insert (nucleotide sensor). The structure of the DOCK9DHR2-Cdc42 complex allowed us to identify a nucleotide sensor within the a10 helix of the DHR2 domain that contributes to release of GDP and then to discharge of the activated GTP-bound Cdc42. The first step of the exchange cycle is to promote nucleotide release by inducing a reduced affinity for nucleotide (Fig. 2). First, the conformational transition of switch 1 of Cdc42 on binding DOCK9DHR2 exposes the nucleotide-binding site. Second, movement of switch 1 is linked to rotation of the P-loop Cys 18 thiol group that disrupts a hydrogen bond with the nucleotide a-phosphate. Third, by intruding into the GTPase nucleotide-binding site, Val 1951 of the a10 insert directly occludes nucleotide-coordinated Mg2+. Because magnesium enhances nucleotide affinity by neutralising the negatively charged phosphate groups, its exclusion profoundly reduces nucleotide affinity. |
To understand the second step of the exchange cycle, loading of GTP-Mg2+ and discharge of the activated Cdc42- GTP-Mg complex, we determined the structure of a GTP-bound ternary complex (data collected at DLS I04). We found that in this structure, Mg2+ bound to GTP caused a displacement of the a10 insert that removes the clamp from switch 1, and induces a set of interdependent conformational changes within other regions of the DOCK9DHR2-Cdc42 interface. This reorganisation of the complex disrupts DOCK9DHR2 contacts to switch 1. In this process activation of Cdc42 is detected by the presence of Mg2+ tightly bound to GTP, triggering displacement of the a10 insert and propagation of conformational changes to the DOCK9DHR2-Cdc42 interface. A structure of GDP bound to the DOCK9DHR2-Cdc42 complex using data collected at DLS I02 showed that because GDP does not induce equivalent conformational changes, GTP may specifically promote discharge of the GTP-bound Cdc42 from DOCK9DHR2 to complete the catalytic exchange reaction. Thus, the a10 insert of DOCK proteins acts as sensor of the GDP and GTP bound states of Rho GTPases to mediate the catalytic cycle of GDP release and subsequent discharge of the GTPbound, activated GTPase.
|
Figure 2. Mechanism of DOCK9DHR2-mediated nucleotide release. Details of conformational | DOCK10 is highly related to DOCK9, but functionally is implicated in mesenchymal tumour migration and metastasis. DOCK10 therefore represents an important anti-cancer drug target. We determined the structure of DOCK10 in complex with Cdc42 using data collected at DLS I02. We extended these studies to investigate DOCK2 which is specific for Rac rather than Cdc42, and implicated as a tumour promoter due to its over-expression in cancer cells. Its major function is to promote metastasis by regulating Rac-dependent cell migration and motility. DOCK2 and DOCK9 share only 18% sequence identity. Major differences are observed at the DOCK2-Rac1 interface from the DOCK10-Cdc42 structure. Native data to 2.6 Å resolution were collected at DLS I02. |
In summary, magnesium exclusion, a critical factor in promoting GDP release, is mediated by a conserved valine residue within this sensor, whereas binding of GTP-Mg2+ to the nucleotide-free complex results in magnesium inducing displacement of the sensor to stimulate discharge of Cdc42- GTP. These studies therefore identified an unusual mechanism of GDP release, and defined the complete GEF catalytic cycle, from GDP dissociation followed by GTP binding and discharge of the activated GTPase.
References
[1] Bos, J.L., Rehmann, H., and Wittinghofer, A. Cell, 129, 865-877 (2007).
[2] Cote, J.F., and Vuori, K. Trends Cell Biol, 17, 383-393 (2007).
[3] Meller, N., Merlot, S., and Guda, C. J, Cell Sci, 118, 4937- 4946 (2005).
[4] Rossman, K.L., Der, C.J., and Sondek, J. Nat Rev Mol Cell Biol, 6, 167-180 (2005).
[5] Yang, J., Zhang, Z., Roe, S.M., Marshall, C.J., and Barford, D. Science, 325, 1398-1402 (2009).
Principal Publications and Authors:
Activation of Rho GTPases by DOCK Exchange Factors Is Mediated by a Nucleotide Sensor. Jing Yang, Ziguo Zhang, Mark Roe, Chris Marshall & David Barford. Science, 325, 1398-1402. (2009).
Funding Acknowledgements
Cancer Research UK Programme Grant Ref C576/A7403. Research carried out at Diamond on I02 and I04, and at the ESRF on ID29.

Diamond Light Source is the UK's national synchrotron science facility, located at the Harwell Science and Innovation Campus in Oxfordshire.
Copyright © 2022 Diamond Light Source
Diamond Light Source Ltd
Diamond House
Harwell Science & Innovation Campus
Didcot
Oxfordshire
OX11 0DE
Diamond Light Source® and the Diamond logo are registered trademarks of Diamond Light Source Ltd
Registered in England and Wales at Diamond House, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 0DE, United Kingdom. Company number: 4375679. VAT number: 287 461 957. Economic Operators Registration and Identification (EORI) number: GB287461957003.

 Science
Science